Michael Kalloniatis and Charles Luu
Retinal Disparity.
When we look at an object with two eyes, we perceive it as singular, like we do other parts of the visual scene stimulating points on our retina that share a common visual direction. These points are termed “retinal corresponding points” and fall on an area called the “horopter”. Points outside the horopter fall on slightly different retinal areas and so do not have the identical visual direction and lead to “retinal disparity”, the basis of our depth discrimination. This retinal image disparity occurs due to the lateral displacement of the eyes. The region in visual space over which we perceive single vision is known as “Panum’s fusional area”, with objects in front and behind this area being in physiological diplopia (i.e. double vision). Our visual system suppresses this diplopia and hence we do not perceive double vision under normal viewing conditions. In order to understand the discussion on the horopter and Panum’s fusional space, the sense of direction will be introduced. Two terms describing direction sense are:
- Oculocentric Visual Direction
- Egocentric Visual Direction
Oculocentric Visual Direction: The visual direction of an object can be represented by a line that joins the object and the fovea called the Principal Visual Direction or visual axis. Based on the principal visual direction, the direction of all other objects in the subjects visual field is determined. This is the called oculocentric visual direction. Therefore, each point of the retina can be considered to have it own sense of direction. For example, when we look at an object, the object is imaged on the fovea. Other objects imaged above the fovea are seen as “below” and those imaged below the fovea are seen as “above”. Visual sense of direction is organised about the fovea. For a given position of the eye, objects having superimposed retinal images will be seen as being in alignment in the visual field, but at a different distance from the eye (figure 1).
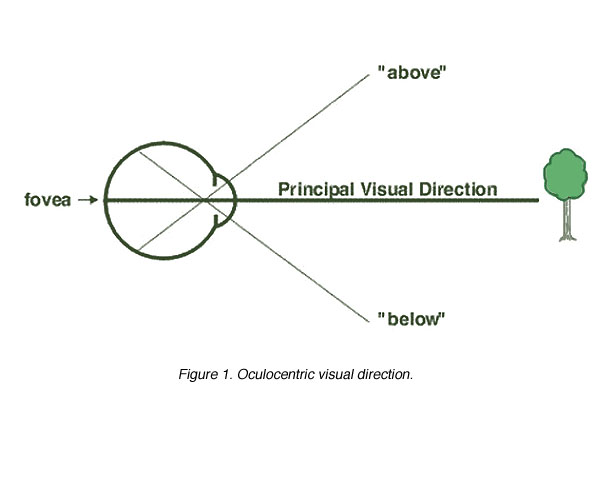
Figure 1. Oculocentric visual direction
Egocentric Visual Direction: Egocentric visual direction refers to the direction of an object in space relative to one self, rather than the eyes. Egocentric direction is determined by retinal position, proprioceptive information about the eye, head and body position and the vestibular apparatus. All this information allows us to determine if a change in retinal position is due to object movement or due to eye or head movement. In figure 2a, a stationary object is imaged on the fovea with the head and the body stationary. When the eye moves, the stationary object is then imaged on a new retinal position. Therefore, oculocentric direction has changed but the egocentric direction has not changed as the object has remained stationary. In another example, the eye tracks a moving object (figure 2b). As the object is imaged on the fovea at all times, the oculocentric direction is the same but the egocentric direction is changing.

In binocular vision, the idea of corresponding retinal points have been used to describe the principle visual direction. Corresponding retinal points are points stimulated on the retina that give rise to the same visual direction. When objects stimulate non-corresponding points, this gives rise to different visual directions. These retinal points are called disparate points. Therefore, corresponding points have the same principle visual direction and non-corresponding points have different visual directions (figure 3).

Figure 3. Corresponding points of the two eyes
As we see the world single and not double, binocular vision can be represented by a single eye, the cyclopean eye. The cyclopean eye is an imaginary eye situated midway between the two eyes (figure 4).
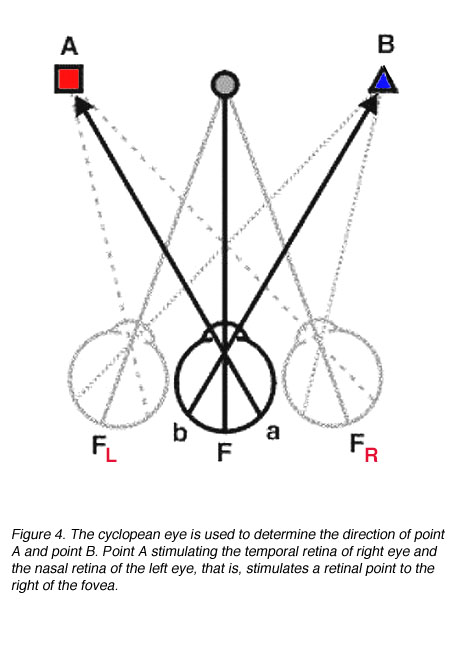
Disparate points give rise to physiological diplopia (double vision). In figure 5, it can be seen that point A stimulates disparate points (non-corresponding retinal points).

Using the cyclopean eye, crossed and uncrossed diplopia can be explored. For an object closer than the fixation point such as point B in figure 6a, crossed diplopia occurs as the point B is imaged on the temporal retina of both eyes. This is termed crossed diplopia because the image in the left eye is seen on the right side. For an object located further than the fixation point, the image of the object falls on the nasal retina of both eyes producing uncrossed diplopia. This is termed uncrossed diplopia because the image in the left eye is seen on the left side (figure 6b).

Figure 6. Demonstrating (a) Crossed and (b) uncrossed diplopia using the cyclopean eye
The principle of the cyclopean eye can be applied to patients with strabismus (a turned eye). Patients with strabismus are usually classified according to the direction of the eye turn. Two common types of strabismus are patients with an esotropia, their eye(s) turned in, and patients with exotropia, their eye(s) turned out. Patients with an exotropia will have crossed diplopia while patients with an esotropia will have uncrossed diplopia (figure 7).

Figure 7. (a) Uncrossed diplopia with an esotropia and (b) crossed diplopia with an exotropia
The Horopter.
Our visual world comprises of multiple points, hence, the need to develop concepts to deal with the whole visual space. This concept is called the horopter. The horopter is the locus of points in space that stimulates corresponding points. That is, a multitude of points in visual space that lead to single vision.
- Vieth-Muller Circle
- Measuring the Horopter
- Relationship of the Horopter to Panum’s fusional area
The Vieth-Muller circle is a theoretical horopter. All points on this circle should stimulate corresponding points on the retina and lead to single vision, provided that the fixation point lies on the centre of the circle and the eyes rotate about its nodal point (instead of their centre of rotation). The Vieth-Muller circle assumes there is angular symmetry of the corresponding points (figure 8).

2) Measuring the horopter
The horopter can be measured through several methods. These methods include:
- Haplopic method
- Nonius method
- Apparent front-parallel plane (AFPP) method
The Nonius and AFPP methods directly determine the longitudinal horopter, whereas the haplopic method does not. Instead, the haplopic method determines the inner and outer boundaries of single binocular vision and the horopter is taken as the midline.
The haplopic method (method of the region of singular binocular vision) is based on the primary definition of corresponding points; retinal points which correspond give rise to identical visual directions and, as a consequence, single vision. Thus if diplopia is observed, disparate points are being stimulated. Therefore the method involves determining the boundaries of single binocular vision (figure 9).

Nonius Method
Since corresponding points give rise to identical visual directions, the position of an object which stimulates a pair of corresponding points can be located if each eye sees a different part of the object. If the two parts are seen in the same direction then the objects are in that position where they stimulate corresponding points. This is the basis of the Nonius method (method of equating visual directions; figure 10).

The Apparent Fronto-Parallel Plane (AFPP) Method
The theory of stereopsis holds that stimulation of disparate points is necessary for the perception of relative depth by stereopsis. If there is no depth difference between an object and the fixation point then they stimulate corresponding points. Thus if the subject is asked to arrange a series of objects so that they appear to be in a fronto-parallel plane (ie. no depth difference between them) then they will lie on the horopter. This is the apparent fronto-parallel plane method (figure 11). Note the change in shape of the horopter at different distances.

Figure 11. The horopter as determined by the apparent fronto-parallel plane method at different distances (25cm, 40cm and 1m). (From Ogle, K. N., Researches in Binocular Vision. London: Saunders, 1950
Relationship of the Horopter to Panum’s Fusional Area
The haplopic method demonstrates the existence of Panum’s fusional area. This concept allows for single binocular vision about the point of fixation even when corresponding retinal points are not being stimulated. An image on the retina of one eye can be fused (and seen as single) with a similar image on the retina of the other eye, even though disparity in the retinal image exists. Panum’s fusional area is needed for stereopsis; if images do not fall in Panum’s area then diplopia results and so Panum’s fusional area defines the zone of stereo vision.
Aniseikonia.
Aniseikonia describes a subject’s spatial perception when there is a difference in retinal image size of the same object between the two eyes. Anisekonia can be investigated by placing an aniseikonic lens placed in front of one eye (to magnify the retinal image in one eye) while plotting the horopter using the AFPP method. When this is performed, the apparent fronto-parallel plane becomes skewed about the fixation point, with the horopter being nearer on the side of the eye having the increased magnification (figure 12). Note that magnification cannot be too large, otherwise, diplopia would result as the two retinal images would fall outside Panum’s fusional area. Aniseikonia identifies reshaping of visual space within Panum’s fusional area.

The importance of these plots with the aniseikonic lens is to demonstrate stable corresponding retinal points. As long as the magnification difference between the two eyes is not too large, fusion will be maintained although spatial distortions will occur. Once the magnification difference exceeds Panum’s fusional space, diplopia will result. Aniseikonic symptoms are a common complaint of patients with unequal refractive errors or large astigmatic corrections. The magical two weeks rule applies, ie, the time taken for sensory adaptation. If symptoms persist, reducing the magnification difference or reducing the correction are two clinical options.
References.
Moses RA and Hart WM (1987) Adler’s Physiology of the eye, Clinical Application, 8th ed. St. Louis: The C. V. Mosby Company.
Ogle KN (1950) Researches in Binocular Vision. London: Saunders.
Schwartz SH (1999)Visual Perception, 2nd ed. Connecticut: Appleton and Lange.
Last Update: June 6, 2007.
The author

Michael Kalloniatis was born in Athens Greece in 1958. He received his optometry degree and Master’s degree from the University of Melbourne. His PhD was awarded from the University of Houston, College of Optometry, for studies investigating colour vision processing in the monkey visual system. Post-doctoral training continued at the University of Texas in Houston with Dr Robert Marc. It was during this period that he developed a keen interest in retinal neurochemistry, but he also maintains an active research laboratory in visual psychophysics focussing on colour vision and visual adaptation. He was a faculty member of the Department of Optometry and Vision Sciences at the University of Melbourne until his recent move to New Zealand. Dr. Kalloniatis is now the Robert G. Leitl Professor of Optometry, Department of Optometry and Vision Science, University of Auckland. e-mail: m.kalloniatis@unsw.edu.au
The author

Charles Luu was born in Can Tho, Vietnam in 1974. He was educated in Melbourne and received his optometry degree from the University of Melbourne in 1996 and proceeded to undertake a clinical residency within the Victorian College of Optometry. During this period, he completed post-graduate training and was awarded the post-graduate diploma in clinical optometry. His areas of expertise include low vision and contact lenses. During his tenure as a staff optometrist, he undertook teaching of optometry students as well as putting together the “Cyclopean Eye”, in collaboration with Dr Michael Kalloniatis. The Cyclopean Eye is a Web based interactive unit used in undergraduate teaching of vision science to optometry students. He is currently in private optometric practice as well as a visiting clinician within the Department of Optometry and Vision Science, University of Melbourne.